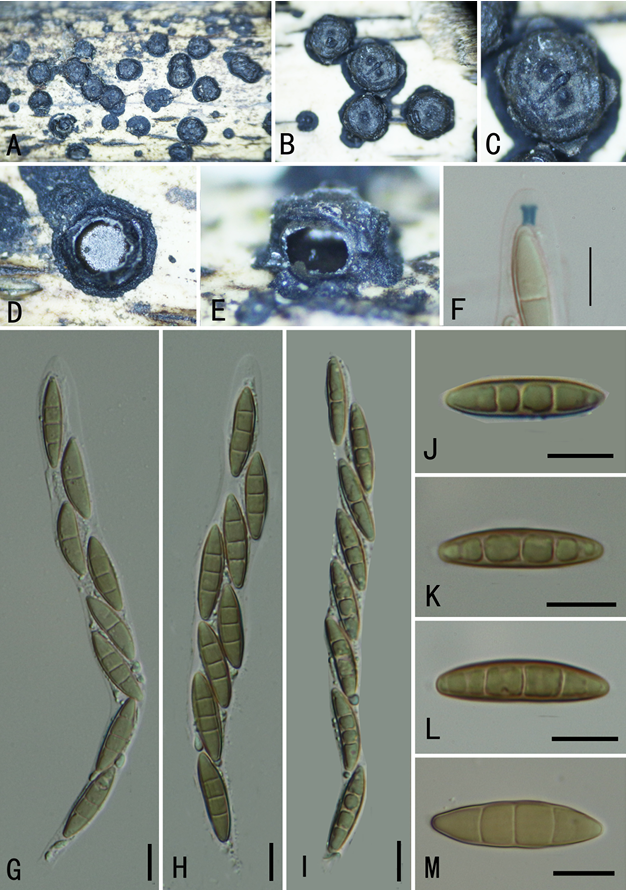
Tremella shuangheensis Q.M. Wang, F.Y. Bai & A.H. Li sp. nov.2020
MycoBank MB828741
Holotype: China, Shuanghe county, Heilongjiang province, obtained from a leaf of an unidentified plant, Aug. 2015, Q.-M. Wang (holotype CGMCC 2.5615T preserved in a metabolically inactive state, ex-type CBS 15561 = SH58A1).
Morphological description
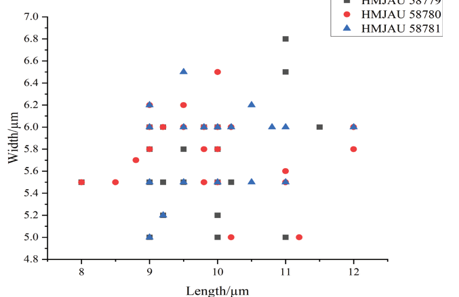
In YM broth, after 7 d at 17 °C, cells are subglobosal and ellipsoidal, 3.2–4.6 × 4.0–5.5 μm and single, budding is polar (Fig. 7G), a sediment is present. After 1 mo at 17 °C, a ring and sediment are present. On YM agar, after 1 mo at 17 °C, the streak culture is yellowish-cream, mucoid, smooth and glistening. The margin is entire. In Dalmau plate culture on corn meal agar, pseudohyphae are not formed. Sexual structures are not observed on YM, PDA, V8 and CM agar. Ballistoconidia are not produced.Physiological and biochemical characteristics: Glucose fermentation is absent. Glucose, galactose (delayed and weak), Lsorbose (weak), sucrose, maltose, cellobiose, trehalose, lactose (delayed), melibiose (delayed), melezitose (delayed and weak), inulin (delayed), soluble starch (delayed and weak), D-xylose, Larabinose (weak), D-arabinose (delayed and weak), D-ribose (delayed and weak), L-rhamnose (delayed and weak), Dglucosamine (delayed and weak), N-Acetyl-D-glucosamine (delayed and weak), ethanol (delayed and weak), glycerol, erythritol, ribitol, galactitol, D-mannitol (delayed and weak), Dglucitol (delayed and weak), Methyl-α-D-glucoside (delayed and weak), salicin (delayed and weak), D-gluconate (delayed and weak), DL-lactate (delayed and weak), succinate (delayed and weak) and myo-inositol (delayed and weak) are assimilated as sole carbon sources. Raffinose, methanol, citrate and hexadecane are not assimilated. Ammonium sulfate, L-lysine, ethylamine hydrochloride and cadaverine dihydrochloride are assimilated. Potassium nitrate and sodium nitrite are not assimilated. Maximum growth temperature is 28 °C. Growth in vitamin-free medium is delayed. Starch-like substances are not produced. Growth on 50 % (w/w) glucose-yeast extract agar is negative. Urease activity is positive. Diazonium Blue B reaction is positive.
Habitat:
Distribution:. China
Reference: Nearly 500 basidiomycetous yeast species were accepted in the latest edition of The Yeasts: A Taxonomic Study published in 2011. However, this number presents only the tip of the iceberg of yeast species diversity in nature. Possibly more than 99 % of yeast species, as is true for many groups of fungi, are yet unknown and await discovery. Over the past two decades nearly 200 unidentified isolates were obtained during a series of environmental surveys of yeasts in phyllosphere and soils, mainly from China. Among these isolates, 107 new species were identified based on the phylogenetic analyses of nuclear ribosomal DNA (rDNA) [D1/D2 domains of the large subunit (LSU), the small subunit (SSU), and the internal transcribed spacer region including the 5.8S rDNA (ITS)] and protein-coding genes [both subunits of DNA polymerase II (RPB1 and RPB2), the translation elongation factor 1-α (TEF1) and the mitochondrial gene cytochrome b (CYTB)], and physiological comparisons. Forty-six of these belong to 16 genera in the Tremellomycetes (Agaricomycotina). The other 61 are distributed in 26 genera in the Pucciniomycotina. Here we circumscribe eight new genera, three new families and two new orders based on the multi-locus phylogenetic analyses combined with the clustering optimisation analysis and the predicted similarity thresholds for yeasts and filamentous fungal delimitation at genus and higher ranks. Additionally, as a result of these analyses, three new combinations are proposed and 66 taxa are validated.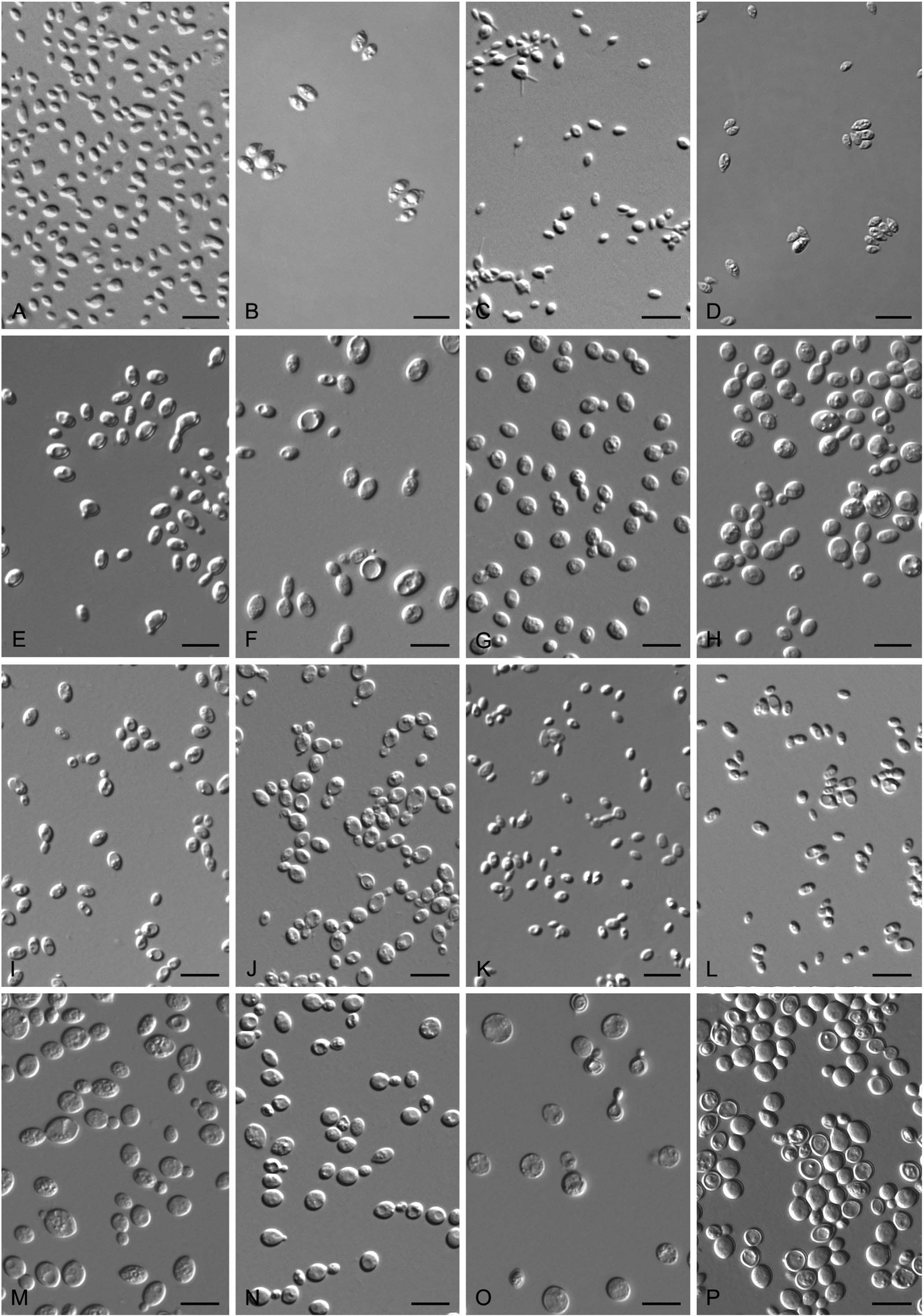
Fig. 7. Vegetative cells grown in YM broth for 5 d at 17 °C and ballistoconidia produced on corn meal agar after 7 d at 17 °C (C, D) Koc. ischaemi CGMCC 2.3565T; ; (G) Tr. shuangheensis CGMCC 2.5615T;